
Abstract
Clearing amyloid-β (Aβ) through immunotherapy is one of the most promising therapeutic approaches to Alzheimer’s disease (AD). Although several monoclonal antibodies against Aβ have been shown to substantially reduce Aβ burden in patients with AD, their effects on improving cognitive function remain marginal. In addition, a significant portion of patients treated with Aβ-targeting antibodies experience brain edema and microhemorrhage associated with antibody-mediated Fc receptor activation in the brain. Here, we develop a phagocytosis inducer for Aβ consisting of a single-chain variable fragment of an Aβ-targeting monoclonal antibody fused with a truncated receptor binding domain of growth arrest-specific 6 (Gas6), a bridging molecule for the clearance of dead cells via TAM (TYRO3, AXL, and MERTK) receptors. This chimeric fusion protein (αAβ–Gas6) selectively eliminates Aβ plaques through TAM receptor-dependent phagocytosis without inducing NF-kB-mediated inflammatory responses or reactive gliosis. Furthermore, αAβ–Gas6 can induce synergistic clearance of Aβ by activating both microglial and astrocytic phagocytosis, resulting in better behavioral outcomes with substantially reduced synapse elimination and microhemorrhage in AD and cerebral amyloid angiopathy model mice compared with Aβ antibody treatment. Our results suggest that αAβ–Gas6 could be a novel immunotherapeutic agent for AD that overcomes the side effects of conventional antibody therapy.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
All primary antibodies used in this study are listed in the Methods. scRNA-seq data were deposited in the GEO database under the accession numbers GSE183755, GSE183757, and GSE183758. Source data are provided with this paper.
References
Busche, M. A. & Hyman, B. T. Synergy between amyloid-β and tau in Alzheimer’s disease. Nat. Neurosci. 23, 1183–1193 (2020).
Duyckaerts, C., Delatour, B. & Potier, M. C. Classification and basic pathology of Alzheimer disease. Acta Neuropathol. 118, 5–36 (2009).
Long, J. M. & Holtzman, D. M. Alzheimer disease: an update on pathobiology and treatment strategies. Cell 179, 312–339 (2019).
Polanco, J. C. et al. Amyloid-β and tau complexity—towards improved biomarkers and targeted therapies. Nat. Rev. Neurol. 14, 22–39 (2018).
Scearce-Levie, K., Sanchez, P. E. & Lewcock, J. W. Leveraging preclinical models for the development of Alzheimer disease therapeutics. Nat. Rev. Drug Discov. 19, 447–462 (2020).
Weninger, S. et al. Active immunotherapy and alternative therapeutic modalities for Alzheimer’s disease. Alzheimers Dement. (N Y) 6, e12090 (2020).
Mintun, M. A. et al. Donanemab in early Alzheimer’s disease. N. Engl. J. Med. 384, 1691–1704 (2021).
Sevigny, J. et al. The antibody aducanumab reduces Aβ plaques in Alzheimer’s disease. Nature 537, 50–56 (2016).
Fuller, J. P., Stavenhagen, J. B. & Teeling, J. L. New roles for Fc receptors in neurodegeneration—the impact on immunotherapy for Alzheimer’s disease. Front. Neurosci. 8, 235 (2014).
Xiong, M. et al. APOE immunotherapy reduces cerebral amyloid angiopathy and amyloid plaques while improving cerebrovascular function. Sci. Transl. Med. https://doi.org/10.1126/scitranslmed.abd7522 (2021).
Wilcock, D. M. et al. Passive immunotherapy against Aβ in aged APP-transgenic mice reverses cognitive deficits and depletes parenchymal amyloid deposits in spite of increased vascular amyloid and microhemorrhage. J. Neuroinflammation 1, 24 (2004).
Klawonn, A. M. et al. Microglial activation elicits a negative affective state through prostaglandin-mediated modulation of striatal neurons. Immunity 54, 225–234 e226 (2021).
Lee, S. H. et al. Antibody-mediated targeting of tau in vivo does not require effector function and microglial engagement. Cell Rep. 16, 1690–1700 (2016).
Leng, F. & Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here. Nat. Rev. Neurol. 17, 157–172 (2021).
Fuchs, Y. & Steller, H. Programmed cell death in animal development and disease. Cell 147, 742–758 (2011).
Doran, A. C., Yurdagul, A. Jr. & Tabas, I. Efferocytosis in health and disease. Nat. Rev. Immunol. 20, 254–267 (2020).
Lemke, G. How macrophages deal with death. Nat. Rev. Immunol. 19, 539–549 (2019).
Dransfield, I., Zagorska, A., Lew, E. D., Michail, K. & Lemke, G. Mer receptor tyrosine kinase mediates both tethering and phagocytosis of apoptotic cells. Cell Death Dis. 6, e1646 (2015).
Fourgeaud, L. et al. TAM receptors regulate multiple features of microglial physiology. Nature 532, 240–244 (2016).
Lemke, G. Phosphatidylserine is the signal for TAM receptors and their ligands. Trends Biochem. Sci. 42, 738–748 (2017).
Lemke, G. Biology of the TAM receptors. Cold Spring Harb. Perspect. Biol. 5, a009076 (2013).
Kojima, Y., Weissman, I. L. & Leeper, N. J. The role of efferocytosis in atherosclerosis. Circulation 135, 476–489 (2017).
van der Meer, J. H., van der Poll, T. & van ’t Veer, C. TAM receptors, Gas6, and protein S: roles in inflammation and hemostasis. Blood 123, 2460–2469 (2014).
Hafizi, S. & Dahlback, B. Gas6 and protein S. Vitamin K-dependent ligands for the Axl receptor tyrosine kinase subfamily. FEBS J. 273, 5231–5244 (2006).
Uehara, H. & Shacter, E. Auto-oxidation and oligomerization of protein S on the apoptotic cell surface is required for Mer tyrosine kinase-mediated phagocytosis of apoptotic cells. J. Immunol. 180, 2522–2530 (2008).
Lew, E. D. et al. Differential TAM receptor-ligand-phospholipid interactions delimit differential TAM bioactivities. eLife https://doi.org/10.7554/eLife.03385 (2014).
Byun, Y. G. & Chung, W. S. A novel in vitro live-imaging assay of astrocyte-mediated phagocytosis using pH indicator-conjugated synaptosomes. J. Vis. Exp. https://doi.org/10.3791/56647 (2018).
Huang, Y. et al. Microglia use TAM receptors to detect and engulf amyloid β plaques. Nat. Immunol. 22, 586–594 (2021).
Linger, R. M., Keating, A. K., Earp, H. S. & Graham, D. K. TAM receptor tyrosine kinases: biologic functions, signaling, and potential therapeutic targeting in human cancer. Adv. Cancer Res. 100, 35–83 (2008).
Tibrewal, N. et al. Autophosphorylation docking site Tyr-867 in Mer receptor tyrosine kinase allows for dissociation of multiple signaling pathways for phagocytosis of apoptotic cells and down-modulation of lipopolysaccharide-inducible NF-κB transcriptional activation. J. Biol. Chem. 283, 3618–3627 (2008).
Rothlin, C. V., Ghosh, S., Zuniga, E. I., Oldstone, M. B. & Lemke, G. TAM receptors are pleiotropic inhibitors of the innate immune response. Cell 131, 1124–1136 (2007).
Sharif, M. N. et al. Twist mediates suppression of inflammation by type I IFNs and Axl. J. Exp. Med. 203, 1891–1901 (2006).
Sosic, D., Richardson, J. A., Yu, K., Ornitz, D. M. & Olson, E. N. Twist regulates cytokine gene expression through a negative feedback loop that represses NF-κB activity. Cell 112, 169–180 (2003).
Liddelow, S. A. et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 541, 481–487 (2017).
Goshi, N., Morgan, R. K., Lein, P. J. & Seker, E. A primary neural cell culture model to study neuron, astrocyte, and microglia interactions in neuroinflammation. J. Neuroinflammation 17, 155 (2020).
Yuan, Q. et al. Intracisternal injection of beta-amyloid seeds promotes cerebral amyloid angiopathy. Brain Behav. Immun. 89, 628–640 (2020).
Keren-Shaul, H. et al. A unique microglia type associated with restricting development of Alzheimer’s disease. Cell 169, 1276–1290 e1217 (2017).
Pinto, B. et al. Rescuing over-activated microglia restores cognitive performance in juvenile animals of the Dp(16) mouse model of Down syndrome. Neuron 108, 887–904.e12 (2020).
Lall, D. et al. C9orf72 deficiency promotes microglial-mediated synaptic loss in aging and amyloid accumulation. Neuron https://doi.org/10.1016/j.neuron.2021.05.020 (2021).
Bolton, J. L. et al. Early stress-induced impaired microglial pruning of excitatory synapses on immature CRH-expressing neurons provokes aberrant adult stress responses. Cell Rep. 38, 110600 (2022).
Diaz-Aparicio, I. et al. Microglia actively remodel adult hippocampal neurogenesis through the phagocytosis secretome. J. Neurosci. 40, 1453–1482 (2020).
Esposito, N. J., Mazzoni, F., Vargas, J. A. & Finnemann, S. C. Diurnal photoreceptor outer segment renewal in mice is independent of Galectin-3. Invest. Ophthalmol. Vis. Sci. 62, 7 (2021).
Feng, W., Yasumura, D., Matthes, M. T., LaVail, M. M. & Vollrath, D. Mertk triggers uptake of photoreceptor outer segments during phagocytosis by cultured retinal pigment epithelial cells. J. Biol. Chem. 277, 17016–17022 (2002).
Stuart, T. et al. Comprehensive integration of single-cell data. Cell 177, 1888–1902 e1821 (2019).
Wlodarczyk, A. et al. A novel microglial subset plays a key role in myelinogenesis in developing brain. EMBO J. 36, 3292–3308 (2017).
Hagemeyer, N. et al. Microglia contribute to normal myelinogenesis and to oligodendrocyte progenitor maintenance during adulthood. Acta Neuropathol. 134, 441–458 (2017).
Chen, Y. & Colonna, M. Microglia in Alzheimer’s disease at single-cell level. Are there common patterns in humans and mice? J. Exp. Med. https://doi.org/10.1084/jem.20202717 (2021).
Dorman, L. C. et al. A type I interferon response defines a conserved microglial state required for effective neuronal phagocytosis. Preprint at bioRxiv https://doi.org/10.1101/2021.04.29.441889 (2022).
Boisvert, M. M., Erikson, G. A., Shokhirev, M. N. & Allen, N. J. The aging astrocyte transcriptome from multiple regions of the mouse brain. Cell Rep. 22, 269–285 (2018).
Habib, N. et al. Disease-associated astrocytes in Alzheimer’s disease and aging. Nat. Neurosci. 23, 701–706 (2020).
Shi, Y. et al. ApoE4 markedly exacerbates tau-mediated neurodegeneration in a mouse model of tauopathy. Nature 549, 523–527 (2017).
Mahan, T. E. et al. Selective reduction of astrocyte apoE3 and apoE4 strongly reduces Aβ accumulation and plaque-related pathology in a mouse model of amyloidosis. Mol. Neurodegener. 17, 13 (2022).
Wang, C. et al. Selective removal of astrocytic APOE4 strongly protects against tau-mediated neurodegeneration and decreases synaptic phagocytosis by microglia. Neuron 109, 1657–1674.e7 (2021).
Trudler, D. et al. Soluble α-synuclein-antibody complexes activate the NLRP3 inflammasome in hiPSC-derived microglia. Proc. Natl Acad. Sci. USA https://doi.org/10.1073/pnas.2025847118 (2021).
Elegheert, J. et al. Lentiviral transduction of mammalian cells for fast, scalable and high-level production of soluble and membrane proteins. Nat. Protoc. 13, 2991–3017 (2018).
Halle, M. et al. Methods to monitor monocytes-mediated amyloid-beta uptake and phagocytosis in the context of adjuvanted immunotherapies. J. Immunol. Methods 424, 64–79 (2015).
Bohlen, C. J. et al. Diverse requirements for microglial survival, specification, and function revealed by defined-medium cultures. Neuron 94, 759–773 e758 (2017).
Lee, J. H. et al. Astrocytes phagocytose adult hippocampal synapses for circuit homeostasis. Nature 590, 612–617 (2021).
Gilles, J. F., Dos Santos, M., Boudier, T., Bolte, S. & Heck, N. DiAna, an ImageJ tool for object-based 3D co-localization and distance analysis. Methods 115, 55–64 (2017).
Acknowledgements
We thank all members of the Chung and Kim laboratory for helpful discussions. This work was supported by the Global Singularity Research Program (Prep Research) (C.H.K.) and an International Collaboration Initiative grant (no. KAIST-N11210255) (W.-S.C.) supported by KAIST, and the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI) and Korea Dementia Research Center (KDRC), funded by the Ministry of Health & Welfare (MOHW) and MSIT, Republic of Korea (HU20C0290) (W.-S.C.). Imaris software and slide scanner were supported by KAIST Bio Core Center.
Author information
Authors and Affiliations
Contributions
W.-S.C. and C.H.K. designed projects and wrote the manuscript. H.J. designed DNA constructs used in this paper; produced all Gas6 fusion proteins; performed in vitro experiments with cell lines including HMC3 and THP-1, western blot, immunoprecipitation and flow cytometry; and analyzed data. S.Y.L. performed in vitro experiments with primary cells and all in vivo experiments, analyzed data, and prepared samples for single-cell RNA sequencing. H.J. and S.L. analyzed bulk RNA-sequencing and single-cell RNA-sequencing data. H.R.C. isolated primary microglia. Y.C. performed cortical tri-culture and analyzed MAP2 fragmentation. M.K. isolated primary microglia and astrocytes and performed qPCR with primary glial cells. H.J. and S.K. generated the Axl-KO HMC3 cell line. Y.L. cloned the aducanumab vector. K.H.H. prepared samples for bulk RNA sequencing and performed FITC-bead uptake assay.
Corresponding authors
Ethics declarations
Competing interests
H.J., S.Y.L., W.-S.C., and C.H.K. have filed a patent application based on this work. The remaining authors declare no competing interests.
Peer review
Peer review information
Nature Medicine thanks the anonymous reviewers for their contribution to the peer review of this work. Primary handling editor: Jerome Staal, in collaboration with the Nature Medicine team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Characterization of TAM receptor expression in HMC3 cells and target binding mediated by Gas6 fusion proteins.
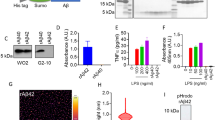
a, mRNA expression levels of TAM and Fc gamma receptors measured by qRT-PCR in HMC3 cells. Mean ± s.e.m. b, Immunoblot analysis for TAM receptor expression using the cell lysates of THP-1 and HMC3 cells. c, Surface expression of TAM and Fc gamma receptors in HMC3 cells analyzed by flow cytometry d, Schematic illustration showing target-specific interaction between beads and AXL receptor induced by Gas6 fusion proteins. e, Gating strategy for flow cytometry analysis to confirm binding of target beads to HMC3 cells. f, Flow cytometry histogram showing the binding of FITC- or oAβ-conjugated beads on HMC3 cells in the presence of different Gas6 fusion proteins. Substitution of the Gla domain with scFv sequence was detected by anti-scFv Fab. g, Flow cytometry analysis showing that Gas6 fusion proteins lost their ability to bind to apoptotic thymocytes generated by treatment with dexamethasone (12.5 μM) for 24 hr. For a, b, representative data from n = 3 biological replicates.
Extended Data Fig. 2 Purification of chimeric Gas6 fusion proteins and characterization of AXL-dependent phagocytosis induced by the fusion proteins.
a, Immunoblot analysis of αAβ-Gas6 and αAβ-Gas6 (E) proteins in HEK293F cell culture supernatants after transduction with lentivirus encoding Gas6 fusion protein genes (NR = Non-Reduced, R = Reduced). b, c, Representative chromatograms of the size exclusion chromatography (SEC) of αAβ-Gas6 (b) and αAβ-Gas6 (E) (c) following FLAG tag affinity chromatography. After αAβ-Gas6 and αAβ-Gas6 (E) were first purified (top), the monomeric form of proteins was collected and subjected to the size exclusion chromatography (bottom). Red arrows indicate the aggregated, dimeric, and monomeric forms of the fusion proteins as determined by molecular weight standards. d, Silver staining showing the monomeric form of αAβ-Gas6 or αAβ-Gas6 after purification. e, Flow cytometry analysis of surface expression of AXL in WT and Axl-KO HMC3 cells. f, Representative kinetic graphs showing the uptake of oAβ-pHrodo in WT and Axl-KO HMC3 cells in the presence of αAβ-Gas6 (5 μg/mL). Unpaired two-tailed t-test at 12 hr after treatment. Mean ± s.e.m.; biological replicates for each group (n = 3). g, Quantification of the phagocytosis of oAβ-pHrodo by HMC3 (after 18-hr incubation) in the presence of αAβ-Gas6 (5 μg/mL) along with a functional blocking antibody for AXL (10 μg/mL). Mean ± s.e.m.; one-way ANOVA followed by Tukey’s multiple comparison test. Representative data from n = 3 (a, d) or 4 (g) biological replicates.
Extended Data Fig. 3 In vitro phagocytosis assays and mechanism studies using Axl-overexpressing THP-1 cells.
a, mRNA expression levels of TAM and Fc gamma receptors measured by qRT-PCR in THP-1 cells. b, Schematic illustration of the generation of THP-1Axl cells expressing both Fc receptors and AXL. c, Flow cytometry analysis showing the expression levels of AXL mutants and d, TAM and Fc gamma receptors in unmodified THP-1 cells. e, Quantification of the phagocytosis of oAβ-pHrodo by THP-1 (left), THP-1Axl (middle), and THP-1Axl△CY (right) in the presence of αAβ-Gas6 (5 μg/mL) for 16 hr. Two-sided unpaired Student’s t-test. f, Representative epi-fluorescence images of THP-1Axl and THP-1Axl△CY showing membrane-bound (orange) and internalized (green) FITC-conjugated beads (8.42 μm, green) after αFITC-Gas6 treatment. Beads number indicates the number of associated beads per 100 cells in each replicate (n = 5). g, Schematic illustration showing the Axl, AxlΔCY, and mutant Axl construct which contains five Y to F mutations h, Representative images of phagocytic events in THP-1 cells expressing different forms of AXL receptor. Scale bar = 200 μm i, Quantification of the phagocytosis events of 5.7 μm beads conjugated with oAβ and pHrodo in the presence of αAβ-Gas6 (5 μg/mL) for 4 hr. j, Immunoblotting showing the forms of Aβ Mo/Di/Trimer (Monomer/Dimer/Trimer) or oligomer. k, Quantitative bar graphs showing the protein levels of pro-inflammatory cytokines (TNF, IL-6, and IL-1β) in THP-1Axl measured by Cytometric Bead Array at 18 hr. l, Luciferase reporter assay for NF-kB transcriptional activity in THP-1Axl. m, Relative mRNA levels of Twist1 (left) and Twist2 (right) by qRT-PCR analysis. The fold change values were normalized to the control group that received only oAβ. Mean ± s.e.m.; one-way (i, k) or two-way (l, m) ANOVA followed by Tukey’s multiple comparisons test. n, Immunoblotting showing the phosphorylation of STAT1 after αAβ-Gas6 (10 μg/mL) treatment of THP-1Axl. For a, e, f, i, k-n, mean ± s.e.m.; representative data from n = 3 biological replicates.
Extended Data Fig. 4 Expression of TAM and Fc gamma receptors in primary rat glial cells.
a, Quantitative bar graphs showing the mRNA levels of Axl, Mer, Cd64, and Cd16 in cultured microglia and astrocytes measured by qRT-PCR at DIV 7. mRNA expression levels were normalized by Axl expression level in each glial cell. Mean ± s.e.m.; b, Immunoblot analysis for TAM receptor expression using the cell lysates of primary microglia and astrocytes. c, Amino acid sequence alignment of LG domains of human, rat, and mouse Gas6 proteins. LG domains of human Gas6 protein shares about 80 percent identity with those of rat or mouse Gas6. The asterisk (*, dark gray), colon (:, gray), and period (., light gray) indicate positions which have a single, fully conserved residue (*), conservation between groups of strongly similar properties (:) or weakly similar properties (.), respectively. d, Immunoprecipitation analysis for tyrosine phosphorylation of AXL in primary microglia or astrocytes. Target cells were incubated with oAβ (2.5 μM) or along with αAβ-Gas6 (4 μg/mL) for 15 min. NT, non-treated. e, Representative Incucyte images of primary microglia (top) and astrocytes (bottom) engulfing oAβ-pHrodo upon treatment of αAβ-Gas6 (5μg/mL) with αTYRO3 (10 μg/mL), αAXL (10μg/mL), or αMERTK (30 μg/mL) antibodies. Scale bar = 100 μm. Mean ± s.e.m. For a, b, d, e, representative data from n = 3 biological replicates.
Extended Data Fig. 5 Validation of protein expression and function in lentiviral system.
a, Coomassie brilliant blue staining of SDS-PAGE gel showing purified Aducanumab (NR = Non-Reduced, R = Reduced). Representative data from n = 3 biological replicates. b, Representative kinetic graphs showing oAβ-pHrodo uptake assays using two different cell lines, THP-1 (FcR-positive) and HMC3 (AXL-positive), which were treated with LV-Aducanumab or LV-αAβ-Gas6, respectively, along with the control virus. One-way ANOVA followed by Tukey’s multiple comparison test. Mean ± s.e.m. c, d, Dot blot assay showing the expression of αAβ-Gas6 (c) or Aducanumab (d) from the lysates of hippocampus 4 weeks after LV-control, LV-αAβ-Gas6, or LV-Aducanumab injection into CA1 region. Anti-DDDDK antibody was used for αAβ-Gas6, while anti-human IgG antibody for Aducanumab. Relative expression value was normalized to the LV-Control group using ImageJ software.
Extended Data Fig. 6 Activation of glial cells in cannula-implanted brains and measurement of plaque burden in CAA-induced brains.
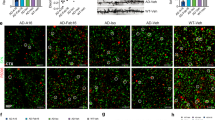
a, Representative confocal z-stack images of MERTK (green), AXL (cyan), GFAP (red), and IBA1 (magenta) in the contralateral (left) and ipsilateral (right) cortex of PBS (top), Aducanumab (middle), and αAβ-Gas6 (bottom) infused 5xFAD mice. White dashed lines indicate the site of cannula implantation. b-g, Representative confocal z-stack images of GFAP (red) and IBA1 (magenta) in the contralateral (b) and ipsilateral (e) hippocampus of PBS (left), Aducanumab (middle), and αAβ-Gas6 (right) infused mice. Quantification of the areas of GFAP (c, f) and IBA1 (d, g) in the imaging fields. n = 6, 8, 6 (c, d, f, g). Scale bars, 100 μm. h, Representative confocal z-stack images of Aβ plaques (Thioflavin-T, green) in contralateral side (top) and ipsilateral side (bottom) of PBS-infused mice. Scale bar = 100 μm. i, j, Quantification bar graphs of the number (left) and area (right) of Thioflavin-T. n = 12, 12. k, l, Representative confocal z-stack images of Aβ plaques (Thioflavin-T, green) in 13m-old APP/PS1 sham control (top) and CAA-induced APP/PS1 (bottom) brains. White arrows indicate CAA. White dashed lines indicate the leptomeningeal surface of cortex. Scale bar = 100 μm (k). Quantification of CAA area in the imaging fields. n = 8, 6 (l). m, n, Quantification bar graphs of total plaque area (m, n) in APP/PS1 (m) and 5xFAD (n). n = 12, 11, 12 (m). n = 12, 12, 12 (n). o, Quantification of insoluble Aβ42 (left) and Aβ40 (right) in PBS and αAβ-Gas6 injected 5xFAD mouse cortices assessed by ELISA. Protein concentrations were normalized to total protein concentrations. n = 11, 8. One-way ANOVA followed by Tukey’s multiple comparison test (c-g, m, n). Two-sided unpaired Student’s t-test (i, j, l, o). n.s., not significant. Mean ± s.e.m. For c-j, n value represents independent experiments from 3 (PBS and αAβ-Gas6) or 4 (Aducanumab) mice. l, o, n value represents the number of mice. For m, n, n values represent independent experiments from 3 mice per group for APP/PS1 (m) and 6 mice per group for 5xFAD (n).
Extended Data Fig. 7 Target specificity and glial responses in LV- αAβ-Gas6 injected mice.
a, b, d, Representative confocal z-stack images of astrocytes (S100β, magenta), microglia (IBA1, red), ZsGreen (green), and DAPI (blue) in the CA1 of LV-Control-injected mouse (a). Orange and cyan arrows indicate ZsGreen+ astrocytes and microglia, respectively. Scale bar = 100 μm. Enlarged images of ZsGreen+ astrocyte (top) and microglia (bottom) are shown in b. Scale bar = 10 μm. Quantification bar graph of ZsGreen+ cell ratio (Microglia / Astrocyte) (d). n = 17, 15, 18. c, e, Representative confocal z-stack images of Aβ plaques (Congo Red, magenta) and ZsGreen (green) in the CA1 of LV-Control (left) and LV-αFITC-Gas6-injected (right) 5xFAD mouse brains. Scale bar = 100 μm (c). Quantification bar graphs of plaque number (left) and area (right) in the CA1 of LV-Control and LV-αFITC-Gas6-injected 5xFAD mouse brains (e). n = 11, 11. f, l, Representative confocal z-stack images of dystrophic neurites (LAMP1, green) and Aβ (6E10, red) in LVs-injected 5xFAD mouse brains. Right panels are enlarged images of orange dotted box in the left and middle panels. Scale bars = 100 μm (f). Quantification bar graph of LAMP1+ dystrophic neurites area (l). n = 40, 40, 32. g, h, m, Representative confocal z-stack images of TAM receptors (g, top: AXL; bottom: MERTK, red), glial cells (h, top: S100β; bottom: IBA1, red), αAβ-Gas6 (HA, green), and Aβ plaques (6E10, blue). Scale bars = 10 μm (g, h). Quantification bar graph of αAβ-Gas6 area associated with glial cells around Aβ plaques (m). n = 7, 7. i, n, o, Representative confocal z-stack images of GFAP (i) in the CA1 of LV-Control, LV-Aducanumab, and LV-αAβ-Gas6-injected mice. Scale bars = 40 μm (i). Quantification of the area (n) and intensity (o) of GFAP in the imaging fields. n = 6, 8, 6. j, k, p, q, Representative confocal z-stack images of plaque-associated microglia (PAM, j) and non-plaque-associated microglia (NPAM, k) in the CA1 of WT (k) and LV-injected 5xFAD mice (j, k) stained with microglia (IBA1, blue), CD68 (red) and Aβ plaques (no. 2454, green). Scale bars = 10 μm. Quantification of the areas of IBA1 and CD68 in PAM (normalized by plaque area) (p) and NPAM (q). n = 19, 21, 21 (p, left); n = 19, 21, 19 (p, right); n = 28, 25, 22, 24 (k, left); n = 19, 24, 22, 23 (k, right). One-way ANOVA followed by Tukey’s multiple comparison test. p, Two-sided unpaired Student’s t-test. Mean ± s.e.m.; n value represents independent experiments from 3 to 5 mice per group (d, e, k-p).
Extended Data Fig. 8 Basal locomotive activities and endogenous functions of TAM receptors are not impaired in LV-αAβ-Gas6 injected mice.
a-c, Quantification bar graphs of total distance (a), velocity (b), and time in center zone (c) in the open field chamber during NOR habituation phase of WT and LV-injected 5xFAD mice. n = 16, 9, 11,10 (a-c). d, f, g, Representative confocal z-stack images of doublecortin (DCX, green) and DAPI (red) in the dentate gyrus (DG) of WT (d, upper) and 5xFAD (d, bottom) mice injected with LVs. Scale bar = 100 μm. Right panels are enlarged images of cyan dotted box in the left images. Scale bar = 10 μm. Quantification of DCX+ cells in the DG of WT (f) and 5xFAD (g) mice. n = 8, 12 (f), n = 11, 12, 12 (g). e, h, i, Representative confocal single plane images of microglia (IBA1, blue), lysosomes (CD68, red), and synapses (top: PSD95; bottom: vGLUT2, green) in the WT injected with LV-Control and LV-αAβ-Gas6 (e). Right panels are 3D reconstructed enlarged images of cyan dotted boxes in left and middle panels, showing engulfed synapses (green, white arrows) in the lysosomes (CD68, red) inside microglia (IBA1, cobalt blue). Scale bars = 3 μm. Quantification bar graphs of area of engulfed synapses (h: PSD95, i: vGLUT2) by microglia normalized by synapse area (h, i). n = 24, 24 (h), n = 24, 23 (i). j, k, Representative confocal z-stack images of photoreceptor outer segment (rhodopsin, green) and nuclear layers (DAPI, blue) in the retina of WT sham control (left) and i.t. injected 5xFAD (middle and right) mice (j). Scale bar = 100 μm. Quantification of length of POS (k). n = 12, 10, 12. a-c, g, k, One-way ANOVA followed by Tukey’s multiple comparison test. Mean ± s.e.m. f, h, i, Two-sided unpaired Student’s t-test. n.s., not significant. Mean ± s.e.m.; a-c, n value represents mice number. For f-i, k, n value represents independent experiments from 3 to 4 mice per group.
Extended Data Fig. 9 scRNA-seq analysis of microglial clusters from LV-injected 5xFAD mice.
a, Violin plots showing the expression levels of representative marker genes for the individual cell types from the 5xFAD hippocampus. b, Cluster fractions of total brain cells of the 5xFAD hippocampus injected with LV-Control, LV-Aducanumab, and LV-αAβ-Gas6. c, Individual UMAP plots of each treatment group. d, Heat map of 3 distinct microglia clusters characterized by suggestive plaque-associated microglia gene sets. e, UMAP clustering of the five microglial signatures (Homeostatic, DAM-like, INF, MHC-II, and Cyc-M) with defined microglial gene modules. The microglial gene signatures were calculated and scored using AddModuleScore function within the Seurat package. f, The bar graph showing the numbers of INF cluster cells in the 5xFAD hippocampus injected with LV-Control, LV-Aducanumab, and LV-αAβ-Gas6. g, Gene Ontology terms showing upregulated cellular pathways in LV-Aducanumab and LV-αAβ-Gas6 injected 5xFAD mice. These pathways were selected by 5 top score of each group after filtering by adjusted p-value <0.05.
Extended Data Fig. 10 Bulk RNA-seq data showing distinct inflammatory responses of THP-1Axl by E2 and αFITC-Gas6 (E)-mediated phagocytosis.
a, Principal Component Analysis (PCA) showing clustering of each group: beads only (Control), beads treated with E2 (E2), and beads treated with αFITC-Gas6 (E) in THP-1Axl. b, Corresponding volcano plots of control versus E2 (left) and control versus αFITC-Gas6 (E) (right). The left plot showed 195 upregulated (log2FC >1, FDR <0.05) and 96 downregulated (log2FC < -1, FDR <0.05) genes, and the right plot showed 157 upregulated (log2FC >1, FDR <0.05) and 104 downregulated (log2FC < -1, FDR <0.05) genes. FC, fold change. c, Scatter plot for the fold change values of shared DEGs in Control versus E2 (y axis) and Control versus αFITC-Gas6 (E) (x axis). d, Gene set enrichment analysis (GSEA) revealed 195 gene-sets and 157 gene-sets enriched for Control versus E2 and Control versus αFITC-Gas6, respectively. Selected pathway filtered by adjusted p-value <0.05. e-f, Gene-set enrichment analysis (GSEA) comparing E2 or αFITC-Gas6 (E). Gene-set terms are indicated as followed; FDR, false discovery rate; NES, normalized enrichment score.
Supplementary information
Supplementary Tables
Supplementary Table 1 (primers) and Supplementary Table 2 (cell lines).
Source data
Source Data Fig. 1
Source data including unprocessed western blots.
Source Data Fig. 2
Source data including unprocessed western blots.
Source Data Fig. 3
Source data.
Source Data Fig. 4
Source data.
Source Data Extended Data Fig. 1
Source data including unprocessed western blots.
Source Data Extended Data Fig. 2
Source data including unprocessed gels.
Source Data Extended Data Fig. 3
Source data including unprocessed western blots.
Source Data Extended Data Fig. 4
Source data including unprocessed western blots.
Source Data Extended Data Fig. 5
Source data.
Source Data Extended Data Fig. 6
Source data.
Source Data Extended Data Fig. 7
Source data.
Source Data Extended Data Fig. 8
Source data.
Source Data Extended Data Fig. 10
Source data.
Rights and permissions
About this article

Cite this article
Jung, H., Lee, S.Y., Lim, S. et al. Anti-inflammatory clearance of amyloid-β by a chimeric Gas6 fusion protein. Nat Med 28, 1802–1812 (2022). https://doi.org/10.1038/s41591-022-01926-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41591-022-01926-9
This article is cited by
-
Brain clearance of protein aggregates: a close-up on astrocytes
Molecular Neurodegeneration (2024)
-
A one-two punch targeting reactive oxygen species and fibril for rescuing Alzheimer’s disease
Nature Communications (2024)
-
Different inflammatory signatures based on CSF biomarkers relate to preserved or diminished brain structure and cognition
Molecular Psychiatry (2024)
-
Fusion protein clears amyloid-β
Nature Reviews Drug Discovery (2022)
